17 Nov Microfluidics for studying the cell polarity
“To migrate efficiently to target locations, cells must integrate receptor inputs while maintaining polarity: a distinct front that leads and a rear that follows. Here we investigate what is necessary to overwrite pre-existing front-rear polarity in neutrophil-like HL60 cells migrating inside straight microfluidic channels. Using subcellular optogenetic receptor activation, we show that receptor inputs can reorient weakly polarized cells, but the rear of strongly polarized cells is refractory to new inputs. Transient stimulation reveals a multi-step repolarization process, confirming that cell rear sensitivity to receptor input is the primary determinant of large-scale directional reversal. We demonstrate that the RhoA/ROCK/myosin II pathway limits the ability of microfluidic receptor inputs to signal to Cdc42 and reorient migrating neutrophils. We discover that by tuning the phosphorylation of myosin regulatory light chain we can modulate the activity and localization of myosin II and thus the amenability of the cell rear to ‘listen’ to receptor inputs and respond to directional reprogramming.”
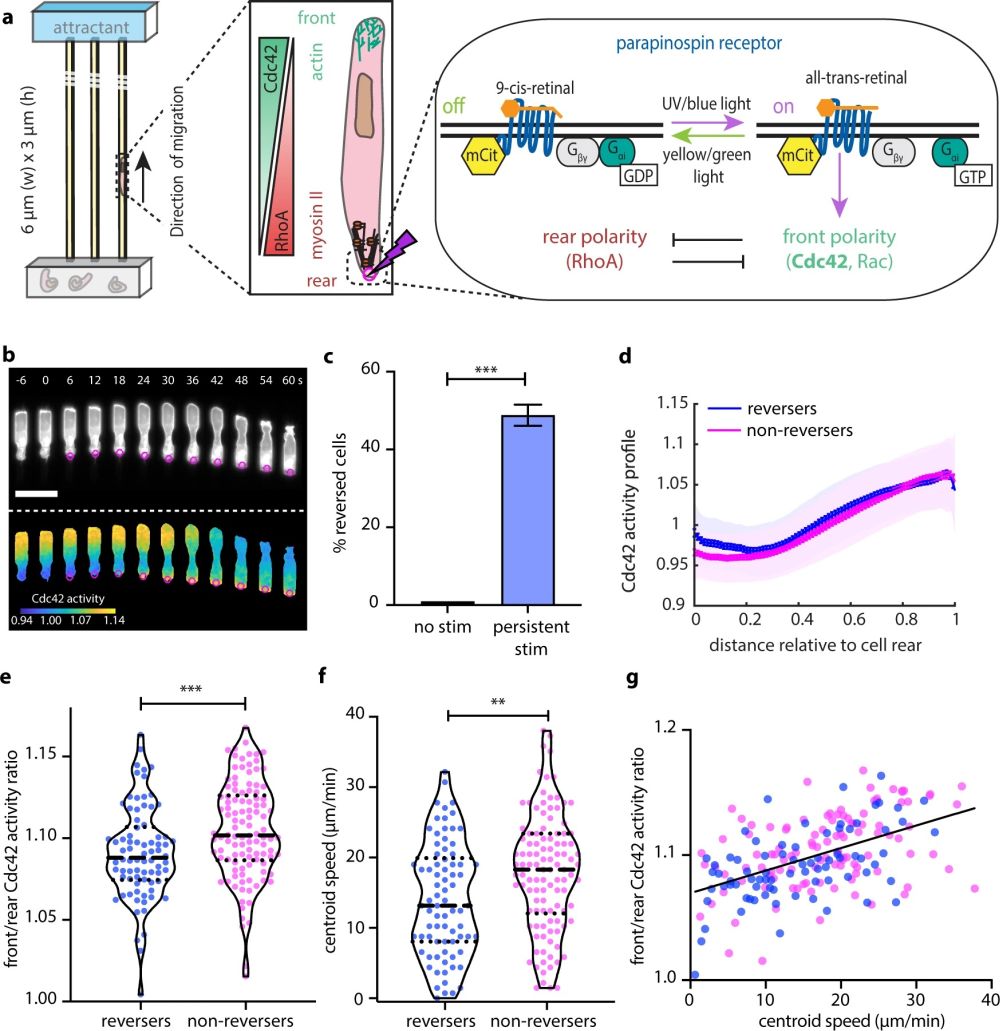
“a Schematic representation of the front–rear polarity reversal assay. HL60 cells expressing parapinopsin, an optogenetic receptor, migrate inside microfluidic devices that harbor straight channels. A heat-inactivated serum gradient is used as an attractant. The opsin can be turned on using UV/blue light and off with yellow/green light, offering complete control of the receptor activity. Blue light stimulation (magenta circle/lighting bolt) at the rear of the cell turns on the receptor. The receptor is coupled to the polarity signal transduction network and activation re-enforces the front module. b Live-cell imaging snapshots of cells expressing parapinopsin and a red/far-red Cdc42 FRET sensor. Cells migrate unperturbed for 60 s prior to initiation of persistent pulsed stimulation (magenta circles) at their rear. Upper and lower panels show the registered sensors (gray scale) and the computed Cdc42 activity, respectively. Images were captured every 3 s and subsampled for illustration purposes. Scale bar: 25 μm. c Box plot of the percentage of cells that reversed with persistent pulsated stimulation (n = 336 cells from 36 independent experiments) vs. non-stimulated control (n = 32 cells from 3 independent experiments). Error bar represents confidence intervals assuming a binomial distribution around the cumulative mean. Two-sided Fisher’s exact test revealed a significant difference between the two conditions (***p < 0.001). d Mean Cdc42 activity profiles for n = 78 reverser (blue) and n = 110 non-reverser (magenta) cells averaging cell profiles over 15 s prior to stimulation (lines: means, shaded regions: SD, error bars: mean value ± SE). e, f Violin plots of mean front/rear Cdc42 activity ratio (e) and of mean centroid speed (f) of n = 78 reverser and n = 110 non-reverser cells, averaging over 15 s prior to stimulation; p-values of two-sided Wilcoxon’s rank-sum test (**p < 0.01, ***p < 0.001). g Linear regression between calculated mean front/rear Cdc42 activity, shown in e, and mean centroid speed, shown in f, prior to stimulation, for n = 188 cells, yields Pearson’s correlation coefficient r = 0.502, p = 2 × 10−13. Source data are provided as a Source Data file, including exact p-values.” Reproduced underCreative Commons Attribution 4.0 International License from Hadjitheodorou, A., Bell, G.R.R., Ellett, F. et al. Nat Commun 12, 6619 (2021).
Figures and the abstract are reproduced from Hadjitheodorou, A., Bell, G.R.R., Ellett, F. et al. Nat Commun 12, 6619 (2021). under Creative Commons Attribution 4.0 International License
Read the original article: Directional reorientation of migrating neutrophils is limited by suppression of receptor input signaling at the cell rear through myosin II activity